
Строение биомембраны. Мембраны, ограничивающие клетки и мембранные органоиды эукариотических клеток, имеют общий химический состав и строение. В их состав входят липиды, белки и углеводы. Липиды мембраны представлены в основном фосфолипидами и холестерином. Большинство белков мембран относится к сложным белкам, например гликопротеинам. Углеводы не встречаются в мембране самостоятельно, они связаны с белками и липидами. Толщина мембран составляет 7-10 нм.
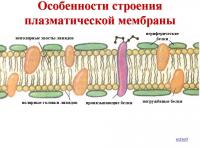
Согласно общепринятой в настоящее время жидкостно-мозаичной модели строения мембран, липиды образуют двойной слой, или липидный бислой, в котором гидрофильные «головки» молекул липидов обращены наружу, а гидрофобные «хвосты» спрятаны вовнутрь мембраны (рис. 2.24). Эти «хвосты» благодаря своей гидрофобности обеспечивают разделение водных фаз внутренней среды клетки и ее окружения. С липидами с помощью различных типов взаимодействия связаны белки. Часть белков расположена на поверхности мембраны. Такие белки называют периферическими, или поверхностными. Другие белки частично или полностью погружены в мембрану - это интегральные, или погруженные белки. Белки мембран выполняют структурную, транспортную, каталитическую, рецеп- торную и другие функции.
Мембраны не похожи на кристаллы, их компоненты постоянно находятся в движении, вследствие чего между молекулами липидов возникают разрывы - поры, через которые в клетку могут попадать или покидать различные вещества.
Биологические мембраны различаются по расположению в клетке, химическому составу и выполняемым функциям. Основные типы мембран - плазматическая и внутренняя.
Плазматическая мембрана (рис. 2.24) содержит около 45% липидов (в т. ч. гликолипидов), 50% белков и 5 % углеводов. Цепочки углеводов, входящих в состав сложных белков-гликопротеинов и сложных липидов-гликолипидов, выступают над поверхностью мембраны. Гликопротеины плазмалеммы чрезвычайно специфичны. Так, например, по ним происходит взаимное узнавание клеток, в том числе сперматозоида и яйцеклетки.
На поверхности животных клеток углеводные цепочки образуют тонкий поверхностный слой - гликокаликс. Он выявлен почти во всех животных клетках, но степень его выраженности неодинакова (10-50 мкм). Гликокаликс обеспечивает непосредственную связь клетки с внешней средой, в нем происходит внеклеточное пищеварение; в гликокаликсе размещены рецепторы. Клетки бактерий, растений и грибов, помимо плазмалеммы, окружены еще и клеточными оболочками.
Внутренние мембраны эукариотических клеток разграничивают различные части клетки, образуя своеобразные «отсеки» - компартменты, что способствует разделению различных процессов обмена веществ и энергии. Они могут различаться по химическому составу и выполняемым функциям, но общий план строения у них сохраняется.
Функции мембран:
1. Ограничивающая. Заключается в том, что они отделяют внутреннее пространство клетки от внешней среды. Мембрана является полупроницаемой, то есть ее свободно преодолевают только те вещества, которые необходимы клетке, при этом существуют механизмы транспорта необходимых веществ.
2. Рецепторная. Связана в первую очередь с восприятием сигналов окружающей среды и передачей этой информации внутрь клетки. За эту функцию отвечают специальные белки-рецепторы. Мембранные белки отвечают еще и за клеточное узнавание по принципу «свой-чужой», а также за образование межклеточных соединений, наиболее изученными из которых являются синапсы нервных клеток.
3. Каталитическая. На мембранах расположены многочисленные ферментные комплексы, вследствие чего на них происходят интенсивные синтетические процессы.
4. Энерготрансформирующая. Связана с образованием энергии, ее запасанием в виде АТФ и расходованием.
5. Компартментализация. Мембраны разграничивают также пространство внутри клетки, разделяя тем самым исходные вещества реакции и ферменты, которые могут осуществлять соответствующие реакции.
6. Образование межклеточных контактов. Несмотря на то, что толщина мембраны настолько мала, что ее невозможно различить невооруженным глазом, она, с одной стороны, служит достаточно надежным барьером для ионов и молекул, в особенности водорастворимых, а с другой - обеспечивает их перенос в клетку и наружу.
Мембранный транспорт. В связи с тем, что клетки как элементарные биологические системы являются открытыми системами, для обеспечения обмена веществ и энергии, поддержания гомеостаза, роста, раздражимости и других процессов требуется перенос веществ через мембрану - мембранный транспорт (рис. 2.25). В настоящее время транспорт веществ через мембрану клетки делят на активный, пассивный, эндо- и экзоцитоз.

Пассивный транспорт - это вид транспорта, который происходит без затраты энергии от большей концентрации к меньшей. Растворимые в липидах небольшие неполярные молекулы (0 2 , С0 2) легко проникают в клетку путем простой диффузии. Нерастворимые же в липидах, в том числе заряженные небольшие частицы, подхватываются белками-пере- носчиками или проходят через специальные каналы (глюкоза, аминокислоты, К + , РО 4 3-). Такой вид пассивного транспорта называется облегченной диффузией. Вода поступает в клеткучерез поры в липидной фазе, а также по специальным каналам, выстланным белками. Транспорт воды через мембрану называется осмосом (рис. 2.26).

Осмос имеет чрезвычайно важное значение в жизни клетки, так как если ее поместить в раствор с более высокой концентрацией солей, чем в клеточном растворе, то вода начнет выходить из клетки, и объем живого содержимого начнет уменьшаться. У животных клеток происходит съеживание клетки в целом, а у растительных - отставание цитоплазмы от клеточной стенки, которое называется плазмолизом (рис. 2.27).

При помещении клетки в менее концентрированный, чем цитоплазма, раствор, транспорт воды происходит в обратном направлении - в клетку. Однако существуют пределы растяжимости цитоплазматической мембраны, и животная клетка в конце концов разрывается, а у растительной этого не позволяет сделать прочная клеточная стенка. Явление заполнения клеточным содержимым всего внутреннего пространства клетки называется деплазмолизом. Внутриклеточную концентрацию солей следует учитывать при приготовлении лекарственных препаратов, особенно для внутривенного введения, так как это может приводить к повреждению клеток крови (для этого используют физиологический раствор с концентрацией 0,9 % хлорида натрия). Это не менее важно при культивировании клеток и тканей, а также органов животных и растений.
Активный транспорт протекает с затратой энергии АТФ от меньшей концентрации вещества к большей. Он осуществляется с помощью специальных белков-насосов. Белки перекачивают через мембрану ионы К + , Na + , Са 2+ и другие, что способствует транспорту важнейших органических веществ, а также возникновению нервных импульсов и т. д.
Эндоцитоз - это активный процесс поглощения веществ клеткой, при котором мембрана образует впячивания, а затем формирует мембранные пузырьки - фагосомы, в которых заключены поглощаемые объекты. Затем с фагосомой сливается первичная лизосома, и образуется вторичная лизосома, или фаголизосома, или пищеварительная вакуоль. Содержимое пузырька расщепляется ферментами лизосом, а продукты расщепления поглощаются и усваиваются клеткой. Непереваренные остатки удаляются из клетки путем экзоцитоза. Различают два основных вида эндоци- тоза: фагоцитоз и пиноцитоз.
Фагоцитоз - это процесс захвата клеточной поверхностью и поглощения клеткой твердых частиц, а пиноцитоз - жидкости. Фагоцитоз протекает в основном в животных клетках (одноклеточные животные, лейкоциты человека), он обеспечивает их питание, а часто и защиту организма (рис. 2.28).

Путем пиноцитоза происходит поглощение белков, комплексов антиген-антитела в процессе иммунных реакций и т. д. Однако путем пиноцитоза или фагоцитоза в клетку также попадают многие вирусы. В клетках растений и грибов фагоцитоз практически невозможен, так они окружены прочными клеточными оболочками.
Экзоцитоз - процесс, обратный эндоцитозу. Таким образом выделяются непереваренные остатки пищи из пищеварительных вакуолей, выводятся необходимые для жизнедеятельности клетки и организма в целом вещества. Например, передача нервных импульсов происходит благодаря выделению посылающим импульс нейроном химических посредников - медиаторов, а в растительных клетках так выделяются вспомогательные углеводы клеточной оболочки.
Клеточные оболочки клеток растений, грибов и бактерий. Снаружи от мембраны клетка может выделять прочный каркас - клеточную оболочку, или клеточную стенку.
У растений основу клеточной оболочки составляет целлюлоза, упакованная в пучки по 50-100 молекул. Промежутки между ними заполняют вода и другие углеводы. Оболочка растительной клетки пронизана каналами - плазмодесмами (рис. 2.29), через которые проходят мембраны эндоплазматической сети.

По плазмодесмам осуществляется транспорт веществ между клетками. Однако транспорт веществ, например воды, может происходить и по самим клеточным стенкам. Со временем в клеточной оболочке растений накапливаются различные вещества, в том числе дубильные или жироподобные, что приводит к одревеснению или опробковению самой клеточной стенки, вытеснению воды и отмиранию клеточного содержимого. Между клеточными стенками соседних клеток растений располагаются желеобразные прокладки - срединные пластинки, которые скрепляют их между собой и цементируют тело растения в целом. Они разрушаются только в процессе созревания плодов и при опадании листьев.
Клеточные стенки клеток грибов образованы хитином - углеводом, содержащим азот. Они достаточно прочны и являются внешним скелетом клетки, но все же, как и у растений, препятствуют фагоцитозу.
У бактерий в состав клеточной стенки входит углевод с фрагментами пептидов - муреин, однако его содержание существенно различается у разных групп бактерий. Кнаружи от клеточной стенки могут выделяться также иные полисахариды, образующие слизистую капсулу, защищающую бактерии от внешних воздействий.
Оболочка определяет форму клетки, служит механической опорой, выполняет защитную функцию, обеспечивает осмотические свойства клетки, ограничивая растяжение живого содержимого и предотвращая разрыв клетки, увеличивающейся вследствие поступления воды. Кроме того, клеточную стенку преодолевают вода и растворенные в ней вещества, прежде чем попасть в цитоплазму или, наоборот, при выходе из нее, при этом по клеточным стенкам вода транспортируется быстрее, чем по цитоплазме.
Клетка — саморегулируемая структурно-функциональная единица тканей и органов. Клеточная теория строения органов и тканей была разработана Шлейденом и Шванном в 1839 г. В дальнейшем с помощью электронной микроскопии и ультрацентрифугирования удалось выяснить строение всех основных органелл животных и растительных клеток (рис. 1).
Рис. 1. Схема строения клетки животных организмов
Главными частями клетки являются цитоплазма и ядро. Каждая клетка окружена очень тонкой мембраной, ограничивающей ее содержимое.
Клеточная мембрана называется плазматической мембраной и характеризуется избирательной проницаемостью. Это свойство позволяет необходимым питательным веществам и химическим элементам проникать внутрь клетки, а излишним продуктам выходить из нее. Плазматическая мембрана состоит из двух слоев липидных молекул с включением в нее специфических белков. Основными липидами мембраны являются фосфолипиды. Они содержат фосфор, полярную головку и два неполярных хвоста из длинноцепочечных жирных кислот. К мембранным липидам относятся холестерин и эфиры холестерина. В соответствии с жидкостно-мозаичной моделью строения, мембраны содержат включения протеиновых и липидных молекул, которые могут перемешаться относительно бислоя. Для каждого типа мембран любой животной клетки характерен свой относительно постоянный липидный состав.
Мембранные белки по структуре подразделяют на два вида: интегральные и периферические. Периферические белки могут удаляться из мембраны без ее разрушения. Имеется четыре типа мембранных белков: транспортные белки, ферменты, рецепторы и структурные белки. Одни мембранные белки обладают ферментативной активностью, другие связывают определенные вещества и способствуют их переносу внутрь клетки. Белки обеспечивают несколько путей передвижения веществ через мембраны: образуют большие поры, состоящие из нескольких белковых субъединиц, которые позволяют перемещаться молекулам воды и ионам между клетками; формируют ионные каналы, специализированные для передвижения ионов некоторых видов через мембрану при определенных условиях. Структурные белки связаны с внутренним липидным слоем и обеспечивают цитоскелет клетки. Цитоскелет придает механическую прочность клеточной оболочке. В различных мембранах на долю белков приходится от 20 до 80% массы. Мембранные белки могут свободно перемещаться в латеральной плоскости.
В мембране присутствуют и углеводы, которые могут ковалентно связываться с липидами или белками. Известно три вида мембранных углеводов: гликолипиды (ганглиозиды), гликопротеиды и протеогликаны. Большинство липидов мембраны находятся в жидком состоянии и обладают определенной текучестью, т.е. способностью перемещаться из одного участка в другой. На внешней стороне мембраны имеются рецепторные участки, связывающие различные гормоны. Другие специфические участки мембраны мог>т распознавать и связывать некоторые чужеродные для данных клеток белки и разнообразные биологически активные соединения.
Внутреннее пространство клетки заполнено цитоплазмой, в которой протекает большинство катализируемых ферментами реакций клеточного метаболизма. Цитоплазма состоит из двух слоев: внутреннего, называемого эндоплазмой, и периферического — эктоплазмы, которая имеет большую вязкость и лишена гранул. В цитоплазме находятся все компоненты клетки или органеллы. Важнейшими из органелл клетки являются — эндоплазматический ретикулум, рибосомы, митохондрии, аппарат Гольджи, лизосомы, микрофиламенты и микротрубочки, пероксисомы.
Эндоплазматический ретикулум представляет собой систему взаимосвязанных каналов и полостей, пронизывающих всю цитоплазму. Он обеспечивает транспорт вешеств из окружающей среды и внутри клеток. Эндоплазматический ретикулум также служит депо для внутриклеточных ионов Са 2+ и служит основным местом синтеза липидов в клетке.
Рибосомы - микроскопические сферические частицы диаметром 10-25 нм. Рибосомы свободно располагаются в цитоплазме или прикрепляются к наружной поверхности мембран эндоплазматической сети и ядерной мембраны. Они взаимодействуют с информационной и транспортной РНК, и в них осуществляется синтез белков. Они синтезируют белки, которые попадают внутрь цистерн или в аппарат Гольджи, и затем выделяются наружу. Рибосомы, свободно располагающиеся в цитоплазме, синтезируют белок для использования самой клеткой, а рибосомы, связанные с эндоплазматическим ретикулумом, производят белок, который выводится из клетки. В рибосомах синтезируются различные функциональные белки: белки-переносчики, ферменты, рецепторы, белки цитоскелета.
Аппарат Гольджи образован системой канальцев, цистерн и пузырьков. Он связан с эндоплазматическим ретикулумом, и поступившие сюда биологически активные вещества хранятся в уплотненном виде в секреторных пузырьках. Последние постоянно отделяются от аппарата Гольджи, транспортируются к клеточной мембране и сливаются с ней, а содержащиеся в пузырьках вещества выводятся из клетки в процессе экзоцитоза.
Лизосомы - окруженные мембраной частицы размером 0,25-0,8 мкм. Они содержат многочисленные ферменты, участвующие в расщеплении белков, полисахаридов, жиров, нуклеиновых кислот, бактерий и клеток.
Пероксисомы сформированы из гладкого эндоплазматического ретикулума, напоминают лизосомы и содержат ферменты, катализирующие разложение пероксида водорода, который расщепляется под влиянием пероксидаз и каталазы.
Митохондрии содержат наружную и внутреннюю мембраны и являются «энергетической станцией» клетки. Митохондрии представляют собой округлые или удлиненные образования с двойной мембраной. Внутренняя мембрана формирует выступающие внутрь митохондрии складки — кристы. В них происходит синтез АТФ, осуществляется окисление субстратов цикла Кребса и множество биохимических реакций. Образованные в митохондриях молекулы АТФ диффундируют во все части клетки. В митохондриях содержится небольшое количество ДНК, РНК, рибосомы, и с их участием происходит обновление и синтез новых митохондрий.
Микрофиламенты представляют собой тонкие белковые нити, состоящие из миозина и актина, и образуют сократительный аппарат клетки. Микрофиламенты участвуют в образовании складок или выпячиваний клеточной мембраны, а также при перемещении различных структур внутри клеток.
Микротрубочки составляют основу цитоскелета и обеспечивают его прочность. Цитоскелет придает клеткам характерные внешний вид и форму, служит местом прикрепления внутриклеточных органелл и различных телец. В нервных клетках пучки микротрубочек участвуют в транспорте веществ из тела клетки к концам аксонов. При их участии осуществляется функционирование митотического веретена во время деления клеток. Они играют роль двигательных элементов в ворсинках и жгутиках у эукариот.
Ядро является основной структурой клетки, участвует в передаче наследственных признаков и в синтезе белков. Ядро окружено ядерной мембраной, содержащей множество ядерных пор, через которые происходит обмен различными веществами между ядром и цитоплазмой. Внутри него находится ядрышко. Установлена важная роль ядрышка в синтезе рибосомной РНК и белков-гистонов. В остальных частях ядра содержится хроматин, состоящий из ДНК, РНК и ряда специфических белков.
Функции клеточной мембраны
В регуляции внутриклеточного и межклеточного обмена важнейшую роль играют клеточные мембраны. Они обладают избирательной проницаемостью. Их специфическое строение позволяет обеспечивать барьерную, транспортную и регуляторную функции.
Барьерная функция проявляется в ограничении проникновения через мембрану растворенных в воде соединений. Мембрана непроницаема для крупных белковых молекул и органических анионов.
Регуляторная функция мембраны состоит в регуляции внутриклеточного метаболизма в ответ на химические, биологические и механические воздействия. Различные воздействия воспринимаются специальными мембранными рецепторами с последующим изменением активности ферментов.
Транспортная функция через биологические мембраны может осуществляться пассивно (диффузия, фильтрация, осмос) или с помощью активного транспорта.
Диффузия - движение газа или растворимого вещества по концентрационному и электрохимическому градиенту. Скорость диффузии зависит от проницаемости клеточной мембраны, а также градиента концентрации для незаряженных частиц, электрического и концентрационного градиентов для заряженных частиц. Простая диффузия происходит через липидный бислой или через каналы. Заряженные частицы движутся согласно электрохимическому градиенту, а незаряженные — химическому градиенту. Например, простой диффузией через липидный слой мембраны проникают кислород, стероидные гормоны, мочевина, спирт и т.д. Через каналы перемещаются различные ионы и частицы. Ионные каналы образованы белками и подразделяются на управляемые и неуправляемые каналы. В зависимости от селективности различают ионоселективные канаты, пропускающие только один ион, и каналы, не обладающие селективностью. Каналы имеют устье и селективный фильтр, а управляемые каналы — и воротный механизм.
Облегченная диффузия - процесс, при котором вещества переносятся через мембрану с помощью специальных мембранных белков- переносчиков. Таким путем в клетку проникают аминокислоты и моносахара. Этот вид транспорта происходит очень быстро.
Осмос - движения воды через мембрану из раствора с более низким в раствор с более высоким осмотическим давлением.
Активный транспорт - перенос веществ против градиента концентрации с помощью транспортных АТФаз (ионных насосов). Этот перенос происходит с затратой энергии.
В большей мере изучены Na + /K + -, Са 2+ - и Н + -насосы. Насосы располагаются на клеточных мембранах.
Разновидностью активного транспорта являются эндоцитоз и экзоцитоз. С помощью этих механизмов транспортируются более крупные вещества (белки, полисахариды, нуклеиновые кислоты), которые не могут переноситься по каналам. Этот транспорт более распространен в эпителиальных клетках кишечника, почечных канальцев, эндотелии сосудов.
При эндоцитозе клеточные мембраны образуют впячивания внутрь клетки, которые отшнуровываясь, превращаются в пузырьки. При экзоцитозе пузырьки с содержимым переносятся к клеточной мембране и сливаются с ней, а содержимое пузырьков выделяется во внеклеточную среду.
Строение и функции клеточной мембраны
Для понимания процессов, обеспечивающих существование электрических потенциалов в живых клетках, прежде всего нужно представлять строение клеточной мембраны и ее свойства.
В настоящее время наибольшим признанием пользуется жидкостно-мозаичная модель мембраны, предложенная С. Сингером и Г. Николсоном в 1972 г. Основу мембраны составляет двойной слой фосфолипидов (бислой), гидрофобные фрагменты молекулы которого погружены в толщу мембраны, а полярные гидрофильные группы ориентированы наружу, т.е. в окружающую водную среду (рис. 2).
Мембранные белки локализованы на поверхности мембраны или могут быть внедрены на различную глубину в гидрофобную зону. Некоторые белки пронизывают мембрану насквозь, и различные гидрофильные группы одного и того же белка обнаруживаются по обе стороны клеточной мембраны. Белки, обнаруженные в плазматической мембране, играют очень важную роль: они участвуют в образовании ионных каналов, играют роль мембранных насосов и переносчиков различных веществ, а также могут выполнять рецептор- ную функцию.
Основные функции клеточной мембраны: барьерная, транспортная, регуляторная, каталитическая.
Барьерная функция заключается в ограничении диффузии через мембрану растворимых в воде соединений, что необходимо для защиты клеток от чужеродных, токсических веществ и сохранения внутри клеток относительного постоянного содержания различных веществ. Так, клеточная мембрана может замедлить диффузию различных веществ в 100 000-10 000 000 раз.

Рис. 2. Трехмерная схема жидкостно-мозаичной модели мембраны Сингера-Николсона
Изображены глобулярные интегральные белки, погруженные в липидный бислой. Часть белков является ионными каналами, другие (гликопротеины) содержат олигосахаридные боковые цепи, участвующие в узнавании клетками друг друга и в межклеточной ткани. Молекулы холестерола вплотную примыкают к фосфолипидным головкам и фиксируют прилегающие участки «хвостов». Внутренние участки хвостов молекулы фосфолипидов не ограничены в своем движении и ответственны за текучесть мембраны (Bretscher, 1985)
В мембране располагаются каналы, через которые проникают ионы. Каналы бывают потенциал зависимыми и потен циалнезависимыми. Потенциалзависимые каналы открываются при изменении разности потенциалов, а потенциалнезависимые (гормонрегулируемые) открываются при взаимодействии рецепторов с веществами. Каналы могут быть открыты или закрыты благодаря воротам. В мембрану встроены два вида ворот: активационные (в глубине канала) и инактивационные (на поверхности канала). Ворота могут находиться в одном из трех состояний:
- открытое состояние (открыты оба вида ворот);
- закрытое состояние (закрыты активационные ворота);
- инактивационное состояние (закрыты инактивационные ворота).
Другой характерной особенностью мембран является способность осуществлять избирательный перенос неорганических ионов, питательных веществ, а также различных продуктов обмена. Различают системы пассивного и активного переноса (транспорта) веществ. Пассивный транспорт осуществляется через ионные каналы с помощью или без помощи белков-переносчиков, а его движущей силой является разность электрохимических потенциалов ионов между внутри- и внеклеточным пространством. Избирательность ионных каналов определяется его геометрическими параметрами и химической природой групп, выстилающих стенки канала и его устье.
В настоящее время наиболее хорошо изучены каналы, обладающие избирательной проницаемостью для ионов Na + , К+ , Са 2+ а также для воды (так называемые аквапорины). Диаметр ионных каналов, по оценкам разных исследований, составляет 0,5-0,7 нм. Пропускная способность каналов может изменяться, через один ионный канал может проходить 10 7 - 10 8 ионов в секунду.
Активный транспорт происходит с затратой энергии и осуществляется так называемыми ионными насосами. Ионные насосы — это молекулярные белковые структуры, встроенные в мембрану и осуществляющие перенос ионов в сторону более высокого электрохимического потенциала.
Работа насосов осуществляется за счет энергии гидролиза АТФ. В настоящее время хорошо изучены Na+/K+ — АТФаза, Са 2+ — АТФаза, Н + — АТФаза, Н + /К + — АТФаза, Mg 2+ — АТФаза, которые обеспечивают перемещение соответственно ионов Na + , К + , Са 2+ , Н+, Mg 2+ изолированно или сопряжено (Na+ и К+; Н+ и К+). Молекулярный механизм активного транспорта до конца не выяснен.
text_fields
text_fields
arrow_upward
Клетки отделены от внутренней среды организма клеточной или плазматической мембраной.
Мембрана обеспечивает:
1) Избирательное проникновение в клетку и из нее молекул и ионов, необходимых для выполнения специфических функций клеток;
2) Избирательный транспорт ионов через мембрану, поддерживая трансмембранную разницу электрического потенциала;
3) Специфику межклеточных контактов.
Благодаря наличию в мембране многочисленных рецепторов, воспринимающих химические сигналы - гормоны, медиаторы и другие биологически активные вещества, она способна изменять метаболическую активность клетки. Мембраны обеспечивают специфику иммунных проявлений, благодаря наличию на них антигенов - структур, вызывающих образование антител, способных специфически связываться с этими антигенами.
Ядро и органеллы клетки также отделены от цитоплазмы мембранами, которые предупреждают свободное движение воды и растворенных в ней веществ из цитоплазмы в них и наоборот. Это создает условия для разделения биохимических процессов, протекающих в различных отсеках (компартментах) внутри клетки.
Структура мембраны клетки
text_fields
text_fields
arrow_upward
Мембрана клетки - эластичная структура, толщиной от 7 до 11 нм (рис.1.1). Она состоит, в основном, из липидоа и белков. От 40 до 90% всех липидов составляют фосфолипиды — фосфатидилхолин, фосфатидилэтаноламин, фосфатидилсерин, сфингомиелин и фосфатидилинозит. Важным компонентом мембраны являются гликолипиды, представленные цереброзидами, сульфатидами, ганглиозидами и холестерином.
Рис. 1.1 Организация мембраны.
Основной структурой мембраны клетки является двойной слой фосфолипидных молекул. За счет гидрофобных взаимодействий углеводные цепочки липидных молекул удерживаются друг возле друга в вытянутом состоянии. Группы же фосфолипидных молекул обоих слоев взаимо действуют с белковыми молекулами, погруженными в липидную мембрану. Благодаря тому, что большинство липидных компонентов бислоя находится в жидком состоянии, мембрана обладает подвижностью, совершает волнообразные движения. Ее участки, а также белки, погруженные в липидный бислой, перемешаются из одной ее части в другую. Подвижность (текучесть) мембран клеток облегчает процессы транспорта веществ через мембрану.
Белки мембраны клеток представлены, в основном, гликопротеинами. Различают:
интегральные белки
, проникающие через всю толщу мембраны и
периферические белки
, прикрепленные только к поверхности мембраны, в основном, к внутренней ее части.
Периферические белки почти все функционируют как энзимы (ацетилхолинестераза, кислая и шелочная фосфатазы и др.). Но некоторые энзимы также представлены интегральными белками - АТФ-аза.
Интегральные белки обеспечивают селективный обмен ионов через каналы мембран между экстрацеллюлярной и интрацеллюлярной жидкостью, а также действуют как белки - переносчики крупных молекул.
Рецепторы и антигены мембраны могут быть представлены как интегральными, так и периферическими белками.
Белки, примыкающие к мембране с цитоплазматической стороны, относятся к цитоскелету клетки . Они могут прикрепляться к мембранным белкам.
Так, белок полосы 3 (номер полосы при электрофорезе белков) эритроцитарных мембран объединяется в ансамбль с другими молекулами цитоскелета - спектрином через низкомолекулярный белок анкирин (рис. 1.2).
Рис. 1.2 Схема расположения белков в примембранном цитоскелете эритроцитов.1 - спектрин; 2 - анкирин; 3 - белок полосы 3; 4 - белок полосы 4,1; 5 - белок полосы 4,9; 6 - олигомер актина; 7 - белок 6; 8 - гпикофорин А; 9 - мембрана.
Спектрин является основным белком цитоскелета, составляющим двумерную сеть, к которой прикрепляется актин.
Актин образует микрофиламенты, представляющие собой сократительный аппарат цитоскелета.
Цитоскелет позволяет клетке проявлять гибкоэластические свойства, обеспечивает дополнительную прочность мембраны.
Большинство интегральных белков - гликопротеины . Их углеводная часть выступает из клеточной мембраны наружу. Многие гликопротеины обладают большим отрицательным зарядом из-за значительного содержания сиаловой кислоты (например, молекула гликофорина). Это обеспечивает поверхности большинства клеток отрицательный заряд, способствуя отталкиванию других отрицательно заряженных объектов. Углеводные выступы гликопротеинов являются носителями антигенов групп крови, других антигенных детерминант клетки, они действуют как рецепторы, связывающие гормоны. Гликопротеины образуют адгезивные молекулы, обуславливающие прикрепление клеток одна к другой, т.е. тесные межклеточные контакты.
Особенности обмена веществ в мембране
text_fields
text_fields
arrow_upward
Мембранные компоненты подвержены многим метаболическим превращениям под влиянием ферментов, расположенных на их мембране или внутри ее. К ним относятся окислительные ферменты, играющие важную роль в модификации гидрофобных элементов мембран - холестерина и др. В мембранах же при активации ферментов - фосфолипаз происходит образование из арахидоновой кислоты биологически активных соединений - простагландинов и их производных. В результате активации метаболизма фосфолипидов в мембране образуются тромбоксаны, лейкотриены, оказывающие мощное воздействие на адгезию тромбоцитов, процесс воспаления и др.
В мембране непрерывно протекают процессы обновления ее компонентов . Так, время жизни мембранных белков колеблется от 2 до 5 дней. Однако в клетке существуют механизмы, обеспечивающие доставку вновь синтезированных молекул белка к мембранным рецепторам, облегчающим встраивание белка в мембрану. «Узнавание» данного рецептора вновь синтезированным белком облегчается образованием сигнального пептида, помогающего найти на мембране рецептор.
Липиды мембраны отличаются также значительной скоростью обмена
, что требует для синтеза этих компонентов мембраны большого количества жирных кислот.
На специфику липидного состава мембран клеток влияют изменения среды обитания человека, характера его питания.
Например, увеличение в пище жирных кислот с ненасыщенными связями увеличивает жидкое состояние липидов мембран клеток различных тканей, приводит к благоприятному для функции мембраны клетки изменению отношения фосфолипидов к сфингомиелинам и липидов к белкам.
Избыток холестерина в мембранах, напротив, увеличивает микровязкость их бислоя фосфолипидных молекул, понижая скорость диффузии некоторых веществ через мембраны клеток.
Пища, обогащенная витаминами А, Е, С, Р улучшает обмен липидов в мембранах эритроцитов, снижает микровязкость мембран. Это повышает деформируемость эритроцитов, облегчает выполнение ими транспортной функции (глава 6).
Дефицит жирных кислот и холестерина в пище нарушает липидный состав и функции мембран клеток.
Например, дефицит жиров нарушает функции мембраны нейтрофилов, что угнетает их способность к движению и фагоцитозу (активный захват и поглощение микроскопических инородных живых объектов и твердых частиц одноклеточными организмами или некоторыми клетками).
В регулировании липидного состава мембран и их проницаемости, регуляции пролиферации клеток важную роль играют активные формы кислорода, образующиеся в клетке сопряженно с нормально протекающими метаболическими реакциями (микросомальным окислением и др.).
Образующиеся активные формы кислорода - супероксидный радикал (О 2), перекись водорода (H 2 О 2) и др. представляют собой чрезвычайно реакционноспособные вещества. Их основным субстратом в реакциях свободнорадикального окисления являются ненасыщенные жирные кислоты, входящие в состав фосфолипидов мембран клетки (так называемые реакции перекисного окисления липидов). Интенсификация этих реакций может вызвать повреждение мембраны клетки, ее барьерной, рецепторной и обменной функций, модификацию молекул нуклеиновых кислот и белков, что ведет к мутациям и инактивации ферментов.
В физиологических условиях интенсификация перекисного окисления липидов регулируется антиоксидазной системой клеток, представленной ферментами, инактивируюшими активные формы кислорода - супероксиддисмутазой, каталазой, пероксидазой и веществами, обладающими антиокислительной активностью - токоферолом (витамин Е), убихиноном и др. Выраженный защитный эффект на мембраны клетки (цитопротекторный эффект) при различных повреждающих воздействиях на организм оказывают простагландины Е и J2, «гася» активацию свободнорадикального окисления. Простагландины защищают слизистую желудка и гепатоциты от химических повреждений, нейроны, клетки нейроглии, кардиомиоциты - от гипоксических повреждений, скелетные мышцы — при тяжелой физической нагрузке. Простагландины, связываясь со специфическими рецепторами на клеточных мембранах стабилизируют бислой последних, уменьшают потерю мембранами фосфолипидов.
Функции рецепторов мембран
text_fields
text_fields
arrow_upward
Химический или механический сигнал вначале воспринимается рецепторами мембраны клетки. Следствием этого является химическая модификация мембранных белков, влекущая активацию «вторичных посредников», обеспечивающих быстрое распространение сигнала в клетке к ее геному, энзимам, сократительным элементам и т.д.
Схематично трансмембранная передача сигнала в клетке может быть представлена следующим образом:
1) Возбужденный воспринятым сигналом рецептор активирует у — белки мембраны клетки. Это происходит при связывании ими гуанозинтрифосфата (ГТФ).
2) Взаимодействие комплекса «ГТФ-у- белки», в свою очередь, активирует фермент - предшественник вторичных посредников, расположенный на внутренней стороне мембраны.
Предшественником одного вторичного посредника - цАМФ, образующегося из АТФ, является фермент аденилатциклаза;
Предшественником других вторичных посредников - инозитолтрифосфата и диацилглицерина, образующихся из фосфатидилинозитол-4,5-дифосфата мембраны, является фермент фосфолипаза С. Кроме того, инозитолтрифосфат мобилизует в клетке еще один вторичный посредник - ионы кальция, участвующие практически во всех регуляторных процессах в клетке. Так, например, образовавшийся инозитолтрифосфат вызывает выброс кальция из эндоплазматического ретикулума и повышение его концентрации в цитоплазме, тем самым включая различные формы клеточного ответа. С помощью инозитолтрифосфата и диацилглицерина регулируется функция гладких мышц и В-клеток поджелудочной железы ацетилхолином, передней доли гипофиза тиреогропин-релизинг фактором, ответ лимфоцитов на антиген и т.д.
В некоторых клетках роль вторичного посредника выполняет цГМФ, образующийся из ГТФ с помощью фермента гуанилатциклазы. Он служит, например, вторичным посредником для натрийуретического гормона в гладких мышцах стенок кровеносных сосудов. цАМФ служит вторичным посредником для многих гормонов - адреналина, эритропоэтина и др. (глава 3).
Цитоплазма — обязательная часть клетки, заключенная между плазматической мембраной и ядром; подразделяется на гиалоплазму (основное вещество цитоплазмы), органоиды (постоянные компоненты цитоплазмы) и включения (временные компоненты цитоплазмы). Химический состав цитоплазмы: основу составляет вода (60-90% всей массы цитоплазмы), различные органические и неорганические соединения. Цитоплазма имеет щелочную реакцию. Характерная особенность цитоплазмы эукариотической клетки — постоянное движение (циклоз ). Оно обнаруживается, прежде всего, по перемещению органоидов клетки, например хлоропластов. Если движение цитоплазмы прекращается, клетка погибает, так как, только находясь в постоянном движении, она может выполнять свои функции.
Гиалоплазма (цитозоль ) представляет собой бесцветный, слизистый, густой и прозрачный коллоидный раствор. Именно в ней протекают все процессы обмена веществ, она обеспечивает взаимосвязь ядра и всех органоидов. В зависимости от преобладания в гиалоплазме жидкой части или крупных молекул, различают две формы гиалоплазмы: золь — более жидкая гиалоплазма и гель — более густая гиалоплазма. Между ними возможны взаимопереходы: гель превращается в золь и наоборот.
Функции цитоплазмы:
- объединение всех компонентов клетки в единую систему,
- среда для прохождения многих биохимических и физиологических процессов,
- среда для существования и функционирования органоидов.
Клеточные оболочки
Клеточные оболочки ограничивают эукариотические клетки. В каждой клеточной оболочке можно выделить как минимум два слоя. Внутренний слой прилегает к цитоплазме и представлен плазматической мембраной (синонимы — плазмалемма, клеточная мембрана, цитоплазматическая мембрана), над которой формируется наружный слой. В животной клетке он тонкий и называется гликокаликсом (образован гликопротеинами, гликолипидами, липопротеинами), в растительной клетке — толстый, называется клеточной стенкой (образован целлюлозой).
Все биологические мембраны имеют общие структурные особенности и свойства. В настоящее время общепринята жидкостно-мозаичная модель строения мембраны . Основу мембраны составляет липидный бислой, образованный в основном фосфолипидами. Фосфолипиды — триглицериды, у которых один остаток жирной кислоты замещен на остаток фосфорной кислоты; участок молекулы, в котором находится остаток фосфорной кислоты, называют гидрофильной головкой, участки, в которых находятся остатки жирных кислот — гидрофобными хвостами. В мембране фосфолипиды располагаются строго упорядоченно: гидрофобные хвосты молекул обращены друг к другу, а гидрофильные головки — наружу, к воде.
Помимо липидов в состав мембраны входят белки (в среднем ≈ 60%). Они определяют большинство специфических функций мембраны (транспорт определенных молекул, катализ реакций, получение и преобразование сигналов из окружающей среды и др.). Различают: 1) периферические белки (расположены на наружной или внутренней поверхности липидного бислоя), 2) полуинтегральные белки (погружены в липидный бислой на различную глубину), 3) интегральные, или трансмембранные, белки (пронизывают мембрану насквозь, контактируя при этом и с наружной, и с внутренней средой клетки). Интегральные белки в ряде случаев называют каналообразующими, или канальными, так как их можно рассматривать как гидрофильные каналы, по которым в клетку проходят полярные молекулы (липидный компонент мембраны их бы не пропустил).

А — гидрофильная головка фосфолипида; В — гидрофобные хвостики фосфолипида; 1 — гидрофобные участки белков Е и F; 2 — гидрофильные участки белка F; 3 — разветвленная олигосахаридная цепь, присоединенная к липиду в молекуле гликолипида (гликолипиды встречаются реже, чем гликопротеины); 4 — разветвленная олигосахаридная цепь, присоединенная к белку в молекуле гликопротеина; 5 — гидрофильный канал (функционирует как пора, через которую могут проходить ионы и некоторые полярные молекулы).
В состав мембраны могут входить углеводы (до 10%). Углеводный компонент мембран представлен олигосахаридными или полисахаридными цепями, связанными с молекулами белков (гликопротеины) или липидов (гликолипиды). В основном углеводы располагаются на наружной поверхности мембраны. Углеводы обеспечивают рецепторные функции мембраны. В животных клетках гликопротеины образуют надмембранный комплекс — гликокаликс, имеющий толщину несколько десятков нанометров. В нем располагаются многие рецепторы клетки, с его помощью происходит адгезия клеток.
Молекулы белков, углеводов и липидов подвижны, способны перемещаться в плоскости мембраны. Толщина плазматической мембраны — примерно 7,5 нм.
Функции мембран
Мембраны выполняют такие функции:
- отделение клеточного содержимого от внешней среды,
- регуляция обмена веществ между клеткой и средой,
- деление клетки на компартаменты («отсеки»),
- место локализации «ферментативных конвейеров»,
- обеспечение связи между клетками в тканях многоклеточных организмов (адгезия),
- распознавание сигналов.
Важнейшее свойство мембран — избирательная проницаемость, т.е. мембраны хорошо проницаемы для одних веществ или молекул и плохо проницаемы (или совсем непроницаемы) для других. Это свойство лежит в основе регуляторной функции мембран, обеспечивающей обмен веществ между клеткой и внешней средой. Процесс прохождения веществ через клеточную мембрану называют транспортом веществ . Различают: 1) пассивный транспорт — процесс прохождения веществ, идущий без затрат энергии; 2) активный транспорт — процесс прохождения веществ, идущий с затратами энергии.
При пассивном транспорте вещества перемещаются из области с более высокой концентрацией в область с более низкой, т.е. по градиенту концентрации. В любом растворе имеются молекулы растворителя и растворенного вещества. Процесс перемещения молекул растворенного вещества называют диффузией, перемещения молекул растворителя — осмосом. Если молекула заряжена, то на ее транспорт влияет и электрический градиент. Поэтому часто говорят об электрохимическом градиенте, объединяя оба градиента вместе. Скорость транспорта зависит от величины градиента.
Можно выделить следующие виды пассивного транспорта: 1) простая диффузия — транспорт веществ непосредственно через липидный бислой (кислород, углекислый газ); 2) диффузия через мембранные каналы — транспорт через каналообразующие белки (Na + , K + , Ca 2+ , Cl -); 3) облегченная диффузия — транспорт веществ с помощью специальных транспортных белков, каждый из которых отвечает за перемещение определенных молекул или групп родственных молекул (глюкоза, аминокислоты, нуклеотиды); 4) осмос — транспорт молекул воды (во всех биологических системах растворителем является именно вода).
Необходимость активного транспорта возникает тогда, когда нужно обеспечить перенос через мембрану молекул против электрохимического градиента. Этот транспорт осуществляется особыми белками-переносчиками, деятельность которых требует затрат энергии. Источником энергии служат молекулы АТФ. К активному транспорту относят: 1) Na + /К + -насос (натрий-калиевый насос), 2) эндоцитоз, 3) экзоцитоз.

Работа Na + /К + -насоса . Для нормального функционирования клетка должна поддерживать определенное соотношение ионов К + и Na + в цитоплазме и во внешней среде. Концентрация К + внутри клетки должна быть значительно выше, чем за ее пределами, а Na + — наоборот. Следует отметить, что Na + и К + могут свободно диффундировать через мембранные поры. Na + /К + -насос противодействует выравниванию концентраций этих ионов и активно перекачивает Na + из клетки, а K + в клетку. Na + /К + -насос представляет собой трансмембранный белок, способный к конформационным изменениям, вследствие чего он может присоединять как K + , так и Na + . Цикл работы Na + /К + -насоса можно разделить на следующие фазы: 1) присоединение Na + с внутренней стороны мембраны, 2) фосфорилирование белка-насоса, 3) высвобождение Na + во внеклеточном пространстве, 4) присоединение K + с внешней стороны мембраны, 5) дефосфорилирование белка-насоса, 6) высвобождение K + во внутриклеточном пространстве. На работу натрий-калиевого насоса тратится почти треть всей энергии, необходимой для жизнедеятельности клетки. За один цикл работы насос выкачивает из клетки 3Na + и закачивает 2К + .
Эндоцитоз — процесс поглощения клеткой крупных частиц и макромолекул. Различают два типа эндоцитоза: 1) фагоцитоз — захват и поглощение крупных частиц (клеток, частей клеток, макромолекул) и 2) пиноцитоз — захват и поглощение жидкого материала (раствор, коллоидный раствор, суспензия). Явление фагоцитоза открыто И.И. Мечниковым в 1882 г. При эндоцитозе плазматическая мембрана образует впячивание, края ее сливаются, и происходит отшнуровывание в цитоплазму структур, отграниченных от цитоплазмы одиночной мембраной. К фагоцитозу способны многие простейшие, некоторые лейкоциты. Пиноцитоз наблюдается в эпителиальных клетках кишечника, в эндотелии кровеносных капилляров.
Экзоцитоз — процесс, обратный эндоцитозу: выведение различных веществ из клетки. При экзоцитозе мембрана пузырька сливается с наружной цитоплазматической мембраной, содержимое везикулы выводится за пределы клетки, а ее мембрана включается в состав наружной цитоплазматической мембраны. Таким способом из клеток желез внутренней секреции выводятся гормоны, у простейших — непереваренные остатки пищи.
Перейти к лекции №5 «Клеточная теория. Типы клеточной организации»
Перейти к лекции №7 «Эукариотическая клетка: строение и функции органоидов»
Краткое описание:
Сазонов В.Ф. 1_1 Строение клеточной мембраны [Электронный ресурс] // Кинезиолог, 2009-2018: [сайт]. Дата обновления: 06.02.2018..__.201_). _Описано строение и функционирование клеточной мембраны (синонимы: плазмалемма, плазмолемма, биомембрана, клеточная оболочка, наружная клеточная оболочка, мембрана клетки, цитоплазматическая мембрана). Эти начальные сведения необходимы как для цитологии, так и для понимания процессов нервной деятельности: нервного возбуждения, торможения, работы синапсов и сенсорных рецепторов.
Клеточная мембрана (плазма лемма или плазмо лемма)
Определение понятия
Клеточная мембрана (синонимы: плазмалемма, плазмолемма, цитоплазматическая мембрана, биомембрана) - это тройная липопротеиновая (т.е. "жиро-белковая") оболочка, отделяющая клетку от окружающей среды и осуществлящая управляемый обмен и связь между клеткой и окружающей её средой.
Главное в этом определении - не то, что мембрана отделяет клетку от среды, а как раз то, что она соединяет клетку с окружающей средой. Мембрана - это активная структура клетки, она постоянно работает.
Биологическая мембрана - это ультратонкая бимолекулярная пленка фосфолипидов, инкрустированная белками и полисахаридами. Эта клеточная структура лежит в основе барьерных, механических и матричных свойств живого организма (Антонов В.Ф., 1996).
Образное представление о мембране
Мне клеточная мембрана представляетсся в виде решетчатого забора с множеством дверей в нём, который окружает некую территорию. Всякая мелкая живность может через этот забор свободно перемещаться туда и обратно. Но более крупные посетители могут входить только через двери, да и то не всякие. У разных посетителей ключи только от своих дверей, и через чужие двери они проходить не могут. Так вот через этот забор постоянно идут потоки посетителей туда и обратно, потому что главная функция мембраны-забора двойная: отделять территорию от окружающего пространства и в то же время соединять её с окружающим пространством. Для этого и существует в заборе множество отверстий и дверей - !
Свойства мембраны
1. Проницаемость.
2. Полупроницаемость (частичная проницаемость).
3. Избирательная (синоним: селективная) проницаемость.
4. Активная проницаемость (синоним: активный транспорт).
5. Управляемая проницаемость.
Как видим, основное свойство мембраны - это её проницаемость по отношению к различным веществам.
6. Фагоцитоц и пиноцитоз.
7. Экзоцитоз.
8. Наличие электрических и химических потенциалов, точнее разности потенциалов между внутренней и наружной сторонами мембраны. Образно можно сказать, что "мембрана превращает клетку в "электрическую батарейку" с помощью управления ионными потоками" . Подробности: .
9. Изменения электрического и химического потенциала.
10. Раздражимость. Специальные молекулярные рецепторы, находящиеся на мембране, могут соединяться с сигнальными (управляющими) веществами, вследствие чего может меняться состояние мембраны и всей клетки. Молекулярные рецепторы запускают биохимические реакции в ответ на соединение с ними лигандов (управляющих веществ). Важно отметить, что сигнальное вещество воздействует на рецептор снаружи, а изменения продолжаются внутри клетки. Получается, что мембрана передала информацию из окружающей среды во внутреннюю среду клетки.
11. Каталитическая ферментативная активность. Ферменты могут быть встроены в мембрану или связаны с её поверхностью (как внутри, так и снаружи клетки), и там они осуществляют свою ферментативную деятельность.
12. Изменение формы поверхности и её площади. Это позволяет мембране образовывать выросты наружу или, наоборот, впячивания внутрь клетки.
13. Способность образовывать контакты с другими клеточными мембранами.
14. Адгезия - способность прилипать к твёрдым поверхностям.
Краткий список свойств мембраны
- Проницаемость.
- Эндоцитоз, экзоцитоз, трансцитоз.
- Потенциалы.
- Раздражимость.
- Ферментная активность.
- Контакты.
- Адгезия.
Функции мембраны
1. Неполная изоляция внутреннего содержимого от внешней среды.
2. Главное в работе клеточной мембраны - это обмен различными веществами между клеткой и межклеточной средой. Этому служит такое свойство мембраны как проницаемость. Кроме того, мембрана регулирует этот обмен за счёт того, что регулирует свою проницаемость.
3. Ещё одна важная функция мембраны - создание разности химических и электрических потенциалов между её внутренней и наружной сторонами. За счёт этого внутри клетка имеет отрицательный электрический потенциал - .
4. Через мембрану осуществляется также информационный обмен между клеткой и окружающей её средой. Специальные молекулярные рецепторы, расположенные на мембране, могут связываться с управляющими веществами (гормонами, медиаторами, модуляторами) и запускать в клетке биохимические реакции, приводящие к различным изменениям в работе клетки или в её структурах.
Видео: Строение мембраны клетки
Видеолекция: Подробно о строении мембраны и транспорте
Строение мембраны
Клеточная мембрана имеет универсальное трёхслойное строение. Её срединный жировой слой является сплошным, а верхний и нижний белковые слои покрывают его в виде мозаики из отдельных белковых участков. Жировой слой является основой, обеспечивающей обособление клетки от окружающей среды, изолирующей её от окружающей среды. Сам по себе он очень плохо пропускает водорастворимые вещества, но легко пропускает жирорастворимые. Поэтому проницаемость мембраны для водорастворимых веществ (например, ионов), приходится обеспечивать специальными белковыми структурами - и .
Ниже представлены микрофотографии реальных клеточных мембран контактирующих клеток, полученные с помощью электронного микроскопа, а также схематический рисунок, показывающий трёхслойность мембраны и мозаичность её белковых слоёв. Для увеличения изображения кликните на него.

Отдельное изображение внутреннего липидного (жирового) слоя клеточной мембраны, пронизанного интегральными встроенными белками. Верхний и нижний белковые слои удалены, чтобы не мешать рассмотрению липидного двойного слоя

Рисунок выше: Неполное схематичное изображение клеточной мембраны (клеточной оболочки), приведённое в Википедии.
Учтите, что наружный и внутренний белковые слои здесь с мембраны сняты, чтобы нам лучше был виден центральный жировой двойной липидный слой. В реальной клеточной мембране сверху и снизу по жировой плёночке (мелкие шарики на рисунке) плавают большие белковые "острова", и мембрана получается более толстой, трёхслойной: белок-жир-белок . Так что она на самом деле похожа на сэндвич из двух белковых "кусков хлеба" с жирным слоем "масла" посередине, т.е. имеет трёхслойное строение, а не двухслойное.
На этом рисунке маленькие голубые и белые шарики соответствуют гидрофильным (смачиваемым) «головкам» липидов, а присоединённые к ним «ниточки» - гидрофобным (несмачиваемым) «хвостам». Из белков показаны только интегральные сквозные мембранные белки (красные глобулы и желтые спирали). Желтые овальные точки внутри мембраны - это молекулы холестерола Желто-зеленые цепочки бусинок на наружной стороне мембраны - цепочки олигосахаридов , формирующие гликокаликс. Гликокаликс - это как бы углеводный ("сахарный") "пушок" на мембране, образованный торчащими из неё длинными углеводно-белковыми молекулами.
Живая - это маленький «белково-жировой мешочек», заполненный полужидким желеобразным содержимым, которое пронизано плёнками и трубочками.
Стенки этого мешочка образованы двойной жировой (липидной) плёночкой, облепленной изнутри и снаружи белками - клеточной мембраной. Поэтому говорят, что мембрана имеет трёхслойное строение : белки-жиры-белки . Внутри клетки также есть множество подобных жировых мембран, которые делят её внутреннее пространство на отсеки. Такими же мембранами окружены клеточные органеллы: ядро, митохондрии, хлоропласты. Так что мембрана - это универсальная молекулярная структура, свойственная всем клеткам и всем живым организмам.

Слева - уже не реальная, а искусственная модель кусочка биологической мембраны: это мгновенный снимок жирового фосфолипидного бислоя (т.е. двойного слоя) в процессе его молекулярно-динамического моделирования. Показана расчётная ячейка модели - 96 молекул ФХ (ф осфатидилх олина) и 2304 молекулы воды, всего 20544 атомов.
Справа - наглядная модель одиночной молекулы того самого липида, из которых как раз и собирается мембранный липидный бислой. Вверху у него гидрофильная (водолюбивая) головка, а снизу - два гидрофобных (боящихся воды) хвостика. У этого липида есть простое название: 1-стероил-2-докозагексаеноил-Sn-глицеро-3-фосфатидилхолин (18:0/22:6(n-3)cis ФХ), но вам нет нужды его запоминать, если вы только не планируете довести своего преподавателя до обморока глубиной своих познаний.
Можно дать и более точное научное определение клетке:
– это ограниченная активной мембраной, упорядоченная, структурированная неоднородная система биополимеров, участвующих в единой совокупности обменных, энергетических и информационных процессов, и также осуществляющих поддержание и воспроизведение всей системы в целом.
Внутри клетка также пронизана мембранами, а между мембранами находится не вода, а вязкий гель/золь изменяемой плотности. Поэтому взаимодействующие молекулы в клетке не плавают свободно, как в пробирке с водным раствором, а в основном сидят (иммобилизованы) на полимерных структурах цитоскелета или внутриклеточных мембранах. И химические реакции поэтому проходят внутри клетки почти как в твердом теле, а не в жидкости. Наружная мембрана, окружающая клетку, также облеплена ферментами и молекулярными рецепторами, что делает её очень активной частью клетки.
Клеточная мембрана (плазмалемма, плазмолемма) - это активная оболочка, отделяющая клетку от окружающей среды и связывающая её с окружающей средой. © Сазонов В.Ф., 2016.
Из этого определения мембраны следует, что она не просто ограничивает клетку, а активно работает , связывая её с окружающей её средой.
Жир, из которого состоят мембраны, - особенный, поэтому его молекулы принято называть не просто жиром, а «липидами», «фосфолипидами», «сфинголипидами» . Мембранная плёночка является двойной, т. е. она состоит из двух плёночек, слипшихся друг с другом. Поэтому в учебниках пишут, что основа клеточной мембраны состоит из двух липидных слоёв (или из "бислоя ", т.е. двойного слоя). У каждого отдельно взятого липидного слоя одна сторона может смачиваться водой, а другая - не может. Так вот, эти плёночки слипаются друг с другом именно своими несмачивающимися сторонами.
Мембрана бактерий
Оболочка прокариотической клетки грамотрицательных бактерий состоит из нескольких слоёв, показанных на рисунке ниже.
Слои оболочки грамотрицательных бактерий:
1. Внутренняя трёхслойная цитоплазматическая мембрана, которая соприкасается с цитоплазмой.
2. Клеточная стенка, которая состоит из муреина.
3. Наружная трёхслойная цитоплазматическая мембрана, которая имеет такую же систему липидов с белковыми комплексами, как и внутренняя мембрана.
Общение грамотрицательных бактериальных клеток с внешним миром через такую сложную трёхступенчатую структуру не даёт им преимущества в выживании в суровых условиях по сравнению с грамположительным бактериями, имеющими менее мощную оболочку. Они точно так же плохо переносят высокие температуры, повышенную кислотность и перепады давления.




Видеолекция: Плазматическая мембрана. Е.В. Шеваль, к.б.н.
Видеолекция: Мембрана как клеточная граница. А. Иляскин
Важность ионных каналов мембраны
Легко понять, что через мембранную жировую плёнку могут проникать в клетку только жирорастворимые вещества. Это жиры, спирты, газы. Например, в эритроцитх прямо через мембрану легко проходят внутрь и наружу кислород и углекислый газ. А вот вода и водорастворимые вещества (например, ионы) просто так через мембрану не могут пройти внутрь любой клетки. Это значит, что для них нужны специальные отверстия. Но если просто сделать отверстие в жировой плёнке, то оно тут же затянется обратно. Что же делать? Выход в природе был найден: надо сделать специальные белковые транспортные структуры и протянуть их сквозь мембрану. Именно так и получаются каналы для пропускания не растворимых в жире веществ - ионные каналы мембраны клетки.
Итак, для придания своей мембране дополнительных свойства проницаемости для полярных молекул (ионов и воды) клетка синтезирует в цитоплазме специальные белки, которые затем встраиваются в мембрану. Они бывают двух типов: белки-транспортёры (например, транспортные АТФазы) и белки-каналоформеры (образователи каналов). Эти белки встраиваются в двойной жировой слой мембраны и формируют транспортные структуры в виде транспортёров или в виде ионных каналов . Через эти транспортные структуры теперь могут проходить различные водорастворимые вещества, которые по-другому проходить сквозь жировую мембранную плёнку не могут.
Вообще, встроенные в мембрану белки ещё называются интегральными , именно потому что они как бы включаются в состав мембраны и пронизывают её насквозь. Другие белки, не интегральные, образуют как бы острова, «плавающие» по поверхности мембраны: либо по её наружной поверхности, либо по внутренней. Ведь всем известно, что жир является хорошей смазкой и скользить по нему получается легко!
Выводы
1. В целом, мембрана получается трёхслойной:
1) наружный слой из белковых «островов»,
2) жировое двухслойное «море» (липидный бислой), т.е. двойная липидная плёнка,
3) внутренний слой из белковых «островов».
Но есть ещё рыхлый наружный слой - гликокаликс, который образуют торчащие из мембраны гликопротеины. Они являются молекулярными рецепторами, с которыми связываются сигнальные управляющие вещества.
2. В мембрану встроены специальные белковые структуры, обеспечивающие её протицаемость для ионов или других веществ. Не надо забывать, что в некоторых местах жировое море пронизано интегральными белками насквозь. И именно интегральные белки образуют специальные транспортные структуры клеточной мембраны (смотрите раздел 1_2 Транспортные механизмы мембраны). Через них вещества попадают внутрь клетки, а также выводятся из клетки наружу.
3. С любой стороны мембраны (наружной и внутренней), а также внутри мембраны могут располагаться белки-ферменты, которые влияют и на состояние самой мембраны и на жизнь всей клетки.
Так что мембрана клетки - это активная изменчивая структура, которая активно работает в интересах всей клетки и связывает её с окружающим миром, а не просто является "защитной оболочкой". Это - самое важное, что надо знать про клеточную мембрану.
В медицине мембранные белки зачастую используются как “мишени” для лекарственных средств. В качестве таких мишеней выступают рецепторы, ионные каналы, ферменты, транспортные системы. В последнее время кроме мембраны мишенью для лекарственных веществ становятся также гены, спрятанные в клеточном ядре.
Видео: Введение в биофизику клеточной мембраны: Структура мембран 1 (Владимиров Ю.А.)
Видео: История, строение и функции клеточной мембраны: Структура мембран 2 (Владимиров Ю.А.)
© 2010-2018 Сазонов В.Ф., © 2010-2016 kineziolog.bodhy.


















